¿Las culturas del maíz?:
Arqueobotánica de las
sociedades hidráulicas
de las tierras bajas
sudamericanas
¿Las culturas del maíz?:
Arqueobotánica de las
sociedades hidráulicas
de las tierras bajas
sudamericanas
J O S É I R I A R T E
UNIVERSITY OF EXETER, UK
R U T H D I C K A U
UNIVERSITY OF EXETER, UK
Iriarte, J. | Dickau, R.
¿LAS CULTURAS DEL MAÍZ? ARQUEOBOTÁNICA DE LAS SOCIEDADES HIDRÁULICAS DE LAS TIERRAS BAJAS SUDAMERICANAS
Resumen
En el presente artículo se presenta una síntesis de los estudios arqueobotánicos realizados en sitios agrícolas de humedales pre-Colombinos de la las tierras bajas sudamericanas. Se resume la evidencia obtenida a través del estudio de macro- y micro-vestigios botánicos recuperados tanto de rasgos agrícolas como de sitios arqueológicos de habitación y sedimentos lacustres asociados a los mismos. El análisis de los datos muestra que una gran diversidad de plantas fueron cultivadas en los campos elevados y drenados y consumidos en los sitios habitacionales asociados incluyendo: a) el maíz, b) los tubérculos y las raíces como la mandioca, la batata dulce, el ñame, posiblemente el ocumo y Cannáceas y Marantáceas, c) vegetales como el maní y el zapallo, d) frutos como palmas, y probablemente el icaco, el maracuyá y la guayaba, e) cultivos industriales como el algodón y posiblemente tinturas como el añil y el achiote, así como se han sugerido f) yerbas medicinales, estimulantes y alucinógenas como la yerba mate, la coca y el yopo. Los resultados muestran que el maíz fue probablemente la planta domestica más importante. Se argumenta que la agricultura intensiva de maíz en campos elevados o drenados junto a otras actividades de subsistencia complementarias fue capaz de sostener poblaciones numerosas de las tierras bajas sudamericanas que transformaron el paisaje a gran escala durante el Holoceno Tardío.
Palabras claves: Zea mays, agricultura pre-Colombina, tierras bajas de Sudamérica
THE MAIZE CULTURES? ARCHAEOBOTANY OF THE HYDRAULIC SOCIETIES OF LOWLAND SOUTH AMERICA
Abstract
In this article, we present a synthesis of archaeobotanical studies from pre-Columbian wetland agricultural sites in lowland South America. We summarize the evidence obtained through the study of macro- and microbotanical remains recovered from agricultural features, as well as archaeological habitation sites and lacustrine sediments associated with them. Analysis of the data show that a broad diversity of plants were cultivated on raised and drained fields, and consumed at associated habitation sites, including: a) maize, b) tubers and root crops such as manioc, sweet potato, yam, and possibly cocoyam, Cannaceae, and Marantaceae taxa, c) vegetables such as peanut and squash, d) fruits such as palms, and probably icaco, passion fruit, and guava, e) industrial cultivars such as cotton and possibly pigment sources like indigo and achiote, and suggested f) medicinal, stimulant, and hallucinogenic herbs such as mate, coca, and yopo. Results show that maize was likely the most important domesticated plant. We argue that intensive maize agriculture on raised or drained fields, along with other complimentary subsistence activities, was able to sustain the large populations in the lowlands of South America who transformed the landscape on a large scale during the late Holocene.
Keywords: Zea mays, pre-Columbian agriculture, South American Lowlands
AS CULTURAS DO MILHO? ARQUEOBOTÂNICA DAS SOCIEDADES HIDRÁULICAS DAS TERRAS BAIXAS SUL-AMERICANAS
Resumo
No presente artigo, se apresenta uma síntese dos estudos arqueobotânicos realizados em sítios agrícolas de zonas úmidas pré-Colombianos das terás baixas sul-americanas. Se resume a evidência obtida através do estudo de macro- e microvestígios botânicos recuperados tanto de feições agrícolas como de sítios arqueológicos de habitação e sedimentos lacustres associados aos mesmos. A análise dos dados mostra que uma grande diversidade de plantas foram cultivadas nos campos elevados e drenados e consumidos nos sítios habitacionais associados incluindo: a) o milho, b) os tubérculos e as raízes como a mandioca, a batata doce, o inhame, possivelmente o taro e Canáceas y Marantáceas, c) vegetais como o amendoim e a abóbora, d) frutos como palmas, y provavelmente a jaboticaba, o maracujá e a goiaba, e) cultivos industriais como o algodão y possivelmente tinturas como o anil e o urucum, assim como se tem sugerido f) verbas medicinais, estimulantes e alucinógenas como a erva mate, a coca e o paricá. Os resultados mostram que o milho foi provavelmente a planta doméstica mais importante. Se argumenta que a agricultura intensiva de milho em campos elevados ou drenados junto a outras atividades de subsistência complementares foi capaz de sustentar populações numerosas nas terras baixas sul-americanas que transformaram a paisagem em grande escala durante o Holoceno tardio.
Palavras-chave: Zea mays, agricultura pré-Colombiana, terras baixas da América do Sul
INTRODUCCIÓN
LA AGRICULTURA DE HUMEDALES EN SUDAMÉRICA
Durante el Holoceno Tardío, las sociedades pre-Colombinas de las tierras bajas sudamericanas realizaron una transformación del paisaje a una escala no registrada anteriormente (Heckenberger y Neves 2009). Las grandes extensiones de sabanas estacionalmente inundables en la periferia de los bosques tropicales Amazónicos, previamente no cultivadas, fueron reclamadas como paisajes agrícolas a través de la construcción de sistemas de campos elevados y sistemas de drenaje con la construcción de canales (Denevan 2001, 2007; Denevan et al. 1987; Erickson 2008; Heckenberger y Neves 2009; Valdéz 2006; Walker 2004).
Siguiendo la terminología utilizada por Turner y Denevan (1985:13-14), en este artículo se utiliza de manera general el término agricultura de humedales para referirnos a la agricultura pre-Colombina de sistemas basados en la elevación y el drenaje de suelos cultivables. Reseñaremos únicamente a estos dos tipos de agricultura de humedales, ya que prácticamente no se cuenta con información arqueobotánica sobre otro tipo de agricultura de humedales que existió en tiempos pre-Colombinos como la denominada “agricultura de recesión” (“recessional agroecosystems”), esto es, el cultivo de una superficie estacionalmente inundable durante la época del año en que el agua se retira y el cual no implica trabajos en tierra. En términos generales, en este articulo se entiende como agricultura de campos elevados a “cualquier superficie de tierra preparada que implique la transferencia y la elevación del suelo por sobre su superficie natural para mejorar las condiciones de cultivo” (Denevan y Turner 1974:24). La agricultura de campos drenados comprende la eliminación del agua de un humedal a través de la excavación de canales, la cual no conlleva a la elevación de una superficie de cultivo (ver también Lombardo et al. 2011).
Los campos elevados o drenados han sido una estrategia de cultivo en zonas estacionalmente inundables brindando a los agricultores pre-Colombinos de: a) mejor drenaje y aireación; b)retención de la humedad, en particular durante la estación seca, haciendo posible la recolección de dos cosechas al año; c) el aumento de la fertilidad a través de la incorporación de los sedimentos orgánicos de los humedales y la fijación del nitrógeno, entre otros y d) posiblemente facilitó la colecta y la limpieza de malezas, entre otros beneficios como la capacidad de realizar piscicultura en los canales (ver Denevan 2001; Renard et al. en prensa b).
Los campos elevados cubren vastas extensiones de las sabanas inundables de Sudamérica. Por ejemplo, en sus estudios pioneros, Denevan (1966) calculó un mínimo de 20,235 hectáreas para Mojos, Bolivia basado en fotografías aéreas. En esta región, las nuevas técnicas disponibles para la identificación de campos elevados combinando imágenes satelitales de alta resolución QuickBird y SIG indican que la densidad de campos elevados parece haber sido ser más considerable de lo que se suponía anteriormente (Lombardo 2010). Solo en un sector de la región de Santa Ana de Yacuma en el noroeste del Beni, Lombardo (2010) estima la presencia de 51,500 hectáreas de campos elevados de tipo plataforma.
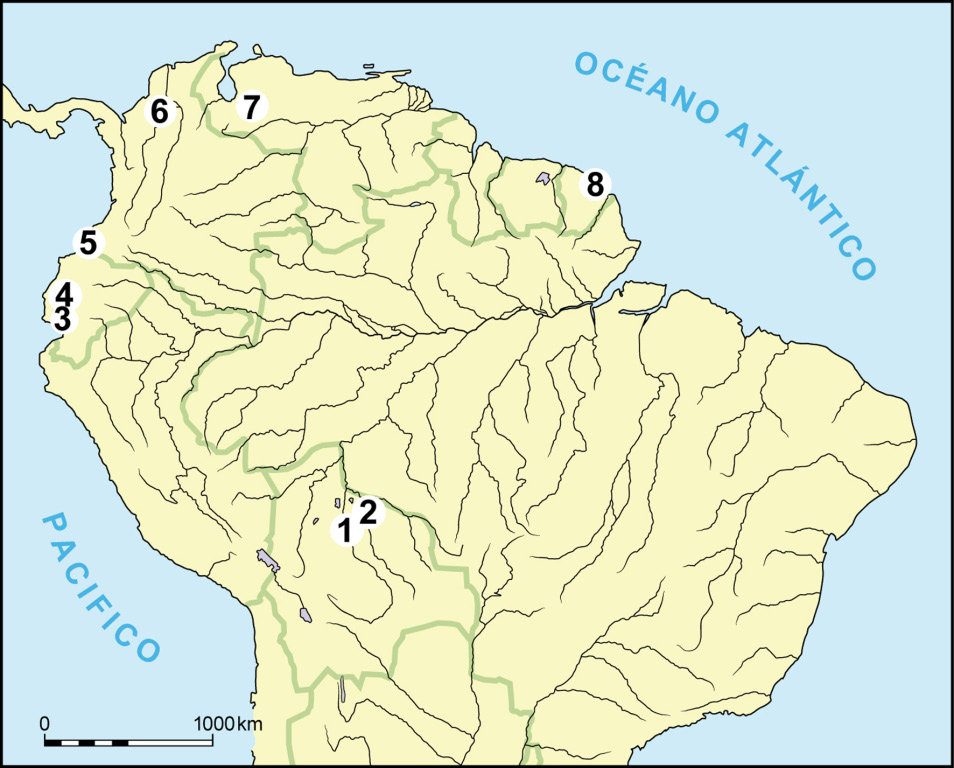
Figura 1 – Mapa de Sudamérica mostrando la ubicación de los sitios con agricultura de humedal mencionados en el texto. 1. Zona central de los Llanos de Mojos al este de la ciudad de Trinidad, Bolivia. 2. Región de Baures, noreste de los Llanos de Mojos, Bolivia. 3. Cuenca baja del Río Guayas, Ecuador. 4. Río Daule, Ecuador. 5. Región de Tolima-Toluca, costa Pacífica de Colombia. 6. Río San Jorge, Colombia. 7. Llanos de Barinas, Venezuela. 8. Costa de la Guyana Francesa.
Existe toda una variedad de campos elevados en las Américas, los cuales pueden llegar hasta 350m de largo, 20m de ancho y más de un metro de altura. Los mismos fueron construidos en una gran diversidad de formas (Denevan 2001:221), posiblemente relacionada con la forma que los agricultores pre-Colombinos manejaron el agua bajo diferentes condiciones de drenaje. Este es el caso de la costa de la Guyana Francesa en donde se encuentran una gran diversidad de campos elevados incluyendo: a) los pequeños montículos redondos (ca.1-1.5m diámetro), conocidos en español como “montones”, los cuales son muy numerosos y cubren completamente algunos parcelas de sabanas inundables (Figura 2a); b) campos elevados más grandes de tamaño mediano, circulares, que pueden tener un diámetro de hasta 5m, los cuales se encuentran principalmente en la pendiente de las elevaciones arenosas de la región y c) los campos alongados (1m altura, 1-5m de ancho y 30-50m de largo), conocidos como “ridged fields” (Denevan 2001) o camellones en español, los cuales generalmente están localizados en las áreas más profundas de las sabanas como en el Bajo Río Mana (Figura 2c)(Rostain 2008, 2010a).
En la mayoría de las regiones donde ocurren los campos elevados o drenados no existe información histórica que ilustre esta práctica agrícola, posiblemente porque muchos de ellos ya habían sido abandonados como producto del Encuentro Colombino. Sin embargo, algunas crónicas europeas tempranas ilustran la práctica de la agricultura de humedales, como es el caso de los indígenas Otomac en Venezuela (Gumilla 1963 [1791]) o los Tainos en Hispaniola (De las Casas 1986 [1560]), los cuales construían pequeños montículos de tierra para cultivar utilizando palas de madera similares a la pala Arauquinoide encontrada en Surinam, la cual data de aprox. 1240 A.D. (Versteeg 2003). Algunos de estos sistemas de campos elevados parecen haber estado en funcionamiento por cientos sino miles de años. Desafortunadamente, la datación directa de los campos elevados es uno de los desafíos más importantes que tiene la arqueología de la agricultura de humedales, aunque se han realizado avances (p. e., McKey et al. 2010; Parsons y Shlemon 1987).
Los experimentos modernos de construcción y cultivo (Arce 1993; Barba et al. 2003; Saavedra 2009; Sanders et al. 1979) tanto como la rehabilitación de campos elevados (Muse y Quintero 1987) muestran que los campos elevados son altamente productivos (al menos en el corto plazo), pudiendo alcanzar una producción de 2 a 5.8 t ha-1 de maíz (Zea mays) y hasta 21 t ha-1 de mandioca (Manihot esculenta).
NUEVAS TÉCNICAS PARA VIEJOS PROBLEMAS
Desde el principio los estudios arqueológicos de las diferentes regiones con agricultura de humedales fueron fuertemente interdisciplinarios. La colaboración conjunta de arqueólogos y geógrafos dio como resultado la aplicación integrada de técnicas como los análisis de fotografía aérea, excavación arqueológica, estudios de suelos, arqueobotánicos, malacológicos, entre otros (p. e., Denevan et al. 1987; Denevan y Zucchi 1978; Stemper 1993; Turner y Harrison 1983). Si bien, muchos de estos estudios han especulado sobre las posibles plantas que se habrían cultivado utilizando agricultura de humedales, mayormente en base a información de las escasas crónicas históricas, pocos datos directos se han obtenido en las tierras bajas sudamericanas hasta ahora.
Nuevos y variados avances metodológicos alcanzados en la arqueobotánica están permitiendo progresar de manera significativa en este tema. Por ejemplo, la aplicación sistemática de análisis de micro-vestigios vegetales está revolucionando nuestro conocimiento sobre las plantas que fueron cultivadas utilizando la agricultura de humedales. Hasta hace muy poco, no éramos capaces de documentar la presencia de raíces de importancia económica, como es el caso de la mandioca o el ñame (Dioscorea sp.) a menos que las mismos fueran recuperadas de ambientes áridos como la costa del Perú (p. e., Ugent et al. 1986) o en ambientes anaeróbicos como las turbas (p. e., Dillehay 1997), en donde las condiciones permitían que los tejidos blandos de estos plantas subterráneas se pudieran preservar. Este es el caso, por ejemplo, de la mandioca, si bien algunas crónicas relatan sobre la importancia de este cultivo en la agricultura de humedales en tiempos históricos (p. e., Castellanos [1522-1607] en Gongard 2006), poca evidencia directa sobre su utilización se había podido recuperar hasta el momento. Este vacío de información ha podido llenarse con el refinamiento de las técnicas especializadas en la recuperación de micro-vestigios botánicos, en particular los estudios de granos de almidón, los cuales están arrojando nueva información sobre la utilización de raíces y tubérculos tanto en las regiones tropicales como templadas de las Américas (p. e., Dickau et al. 2007; Duncan et al. 2009; Chandler-Ezell et al. 2006; Iriarte et al. 2004; Piperno et al. 2000).

Figura 2 – Campos elevados de Sudamérica. A y C. Campos elevados de la Guyana Francesa (Fotos: S. Rostain). B. Campos elevados de la región de San Borja, Llanos de Mojos, Bolivia (Google Earth); D. Plataformas agrícolas de la región de Santa Ana de Yacuma, Llanos de Mojos, Bolivia.
La extracción de granos de almidón y fitolitos de los residuos de los instrumentos para procesar, contener o almacenar plantas (como ser las manos de moler, los ralladores o los coladores, las vasijas cerámicas, entre otros) está permitiendo documentar el rol de estos cultivos en la dieta de las poblaciones pre-Colombinas así como entender las funciones de estos instrumentos (Dickau et al. 2012; McKey et al. 2010). Este último aspecto está obligándonos a revisar las interpretaciones tradicionales, basadas en crónicas históricas y analogías etnográficas, de los instrumentos para procesar vegetales como los dientes de los ralladores de mandioca de cuarzo (p. e., Dickau et al. 2012; Perry 2004, 2005).
Los análisis de fitolitos también están contribuyendo de manera significativa a documentar los cultivos que eran plantados en los campos elevados y/o drenados. Los fitolitos nos brindan varias ventajas para investigar que plantas eran cultivadas en estos sistemas agrícolas: a) debido a su naturaleza silícea (son probablemente el fósil vegetal más robusto que existe), estos se preservan en los suelos de las sabanas tropicales a diferencia de otros vestigios botánicos (Pearsall 1995), b) los mismo se depositan in situ, lo cual resulta muy ventajoso para estudiar qué cultivos fueron plantados localmente en los campos elevados; y (c) algunas plantas como el maíz producen fitolitos diagnósticos en diferentes partes de su estructura, lo que permite identificar diferentes áreas de actividad (p. e., Iriarte 2003; Mulholland 1993) o sugerir la incorporación de abono verde a los campos elevados como una técnica para aumentar la fertilidad (Iriarte et al. 2010). Como nota de precaución, se debe señalar que no todas las plantas domesticas producen fitolitos diagnósticos y por lo tanto, el registro de fitolitos va a ser “silencioso” a la hora de registrar algunas de ellas, como es el caso de los tubérculos como la mandioca, la batata dulce y el ñame (ver Piperno 2006:7, Tabla 1.1). Por lo tanto, se debe complementar el análisis de fitolitos con otras clases de vestigios botánicos para obtener una reconstrucción completa.
Asimismo, el aumento de las colecciones de referencia y el reconocimiento de rasgos micro-morfológicos junto a la aplicación de técnicas de análisis estadísticos multivariados están permitiendo la distinción de fitolitos, granos de almidón y polen a niveles taxonómicos más específicos, así como la diferenciación de especies domesticas de silvestres (p. e., Burn y Mayle 2008; Pearsall et al. 2003; Piperno 2006; Piperno y Stothert 2003). De la misma forma, la aplicación en tándem de fitolitos, polen y carbón para analizar columnas de sedimento recuperadas de lagos y humedales están facilitando una reconstrucción de la historia de la vegetación y de los incendios más completa y precisa (p. e., Bush et al. 1992; Denham et al. 2003; Iriarte 2006; Kealhofer y Penny 1998; Piperno y Jones 2003; Iriarte et al. 2012). Asimismo, la combinación de análisis de fitolitos e isótopos de carbón permiten realizar reconstrucciones más detalladas de los paisajes antiguos (p. e., Alexandre et al. 1999; Iriarte et al. 2010; Renard et al. en prensa a).
Otro de los problemas que enfrenta la arqueobotánica es la dificultad de recuperar granos de polen en sedimentos lagunares y de humedales de plantas domesticas utilizando las técnicas de conteo tradicionales. Los mismos quedan por lo general sub-representados debido a que producen polen de gran tamaño y el cual, por lo general, tiene muy poca dispersión. Se puede citar el caso del polen de maíz que se propaga muy poco de su planta de origen debido a su gran tamaño (entre 55-120µm) (Herrera y Urrego 1996; Jarosz et al. 2003). Lo mismo sucede con otros cultivos como la batata dulce (Ipomea batatas), el zapallo (Cucurbita spp.), y la mandioca, las cuales producen grandes granos de polen (80-250 µm) y debido a que son polinizados por insectos, los mismos arrojan muy pocas cantidades de polen (Hurd et al. 1971; Real 1981; Rival y McKey 2008; Rogers 1965). Estos factores se combinan para que sea muy difícil recuperar polen de estos cultivos en sedimentos lagunares o de humedales utilizando las técnicas tradicionales. Afortunadamente, el desarrollo de metodologías simples de cernir y concentrar los granos de polen por tamaño está facilitando la recuperación más efectiva de granos de plantas domesticadas de granos grandes y dispersión pobre como el maíz, la mandioca, y la batata dulce, entre otras (ver detalles en Whitney et al. en prensa).
RESULTADOS
La información resumida en este artículo comprende datos arqueobotánicos de campos agrícolas elevados/drenados, sitios habitacionales asociados a los mismos y sedimentos lagunares de las siguientes regiones de Sudamérica: Llanos de Mojos, Beni, incluyendo 1) los montículos habitacionales artificiales conocidos como “lomas de habitación” ubicados al este de Trinidad, de los cuales fueron analizados Loma Salvatierra y Loma Mendoza (Bruno 2010; Dickau et al. 2012; Lombardo y Prümers 2010; Prümers 2009) y 2) una zanja circular (“ring ditch”) de la región del Baures, denominada Granja del Padre (Dickau et al. 2012; Prümers 2010; ver también Erickson 2008); 3) la cuenca del Río Guayas, Ecuador (Pearsall 1987); 4) el Río Daule, Ecuador (Stemper 1993); 5) la depresión Momposina en el Bajo Río San Jorge, Colombia (Herrera y Berrio 1998; Plazas y Falchetti 1987; Sneider Rojas y Montejo 2006); 6) la región de Tolima-Toluca, costa Pacífica de Colombia (Patiño 2006); 7) Barinas, Gran Sabana de Venezuela (Redmond y Spencer 2007; Spencer y Redmond 1998; Spencer et al. 1994) y 8) la costa de la Guyana Francesa incluyendo los sitios de el Bajo Río Mana, Grand Macoua, Sable Blanc, K-VIII y Bois Diable (Iriarte et al. 2010; McKey et al. 2010; Rostain 2008, 2010a; Iriarte et al. 2010, 2012; Mckey et al. 2010) (Figura 1).
Tabla 1.
Plantas de importancia económica recuperadas en sitios con agricultura de humedal y sitios habitacionales y sedimentos lagunares asociados en Sudamérica
|
Sitio/Contexto |
Zea mays (maiz) |
Manihot esculenta (mandioca) |
Dioscoreaceae, Dioscorea spp. (ñame) |
Convolvulaceae, cf. Ipomoea (batata dulce) |
Araceae, cf. Xanthosoma sagittifolium (ocumo) |
Marantaceae |
Marantaceae, cf. Maranta arundinacea (arrurruz) |
Cannaceae, Canna edulis (achira) |
Heliconia sp. |
Cyperaceae |
Cucurbita spp. (zapallo) |
Cucurbitaceae, cf. Cucurbita maxima, C. mixta (zapallo) |
Arecaceae (palmeras) |
Arecaceae, cf. Acrocomia sclerocarpa (corozo) |
Astrocaryum standleyanum |
Phytelephas seemannii (tagua) |
Fabaceae |
Arachis hypogaeae (maní) |
Fabaceae, cf. Piptadenia peregrina (yopo) |
Fabaceae, cf. Herpalyce |
Fabaceae, cf. Indigofera tinctoria (añil) |
Aquifoliaceae, cf. Ilex paraguariensis (yerba mate) |
Bixaceae, cf. Bixa orellana (achiote) |
Amaranthaceae/Caryophyllaceae |
Fuente |
|
Llanos de Moxos, Bolivia Campos elevados El Villar, Santa Fe |
p1 |
p1 |
p1 |
Erickson 1995 |
|||||||||||||||||||||
|
Sitios habitación |
|||||||||||||||||||||||||
|
Loma Mendoza |
fgm1,2 |
g2 |
f2 |
f1 |
f1,2 |
f1,2 |
Dickau et al. 2012 |
||||||||||||||||||
|
Loma Salvatierra |
fg2 |
g2 |
g2 |
f1,2 |
m1 |
fm1,2 |
m1 |
m1 |
Bruno 2010; Dickau et al. 2012 |
||||||||||||||||
|
Bella Vista |
fg2 |
g2 |
f1,2 |
f1 |
f1,2 |
Dickau et al. 2012 |
|||||||||||||||||||
|
Tumaco-Tolita, Colombia |
|||||||||||||||||||||||||
|
Campos elevados |
|||||||||||||||||||||||||
|
Las Tres Marias |
f1 |
f1 |
m1 |
m1 |
Patiño 2006 |
||||||||||||||||||||
|
Bajo Río San Jorge, Colombia |
|||||||||||||||||||||||||
|
Sedimento lacunar |
p |
p |
p |
Herrera y Berrio 1998 |
|||||||||||||||||||||
|
Peñon del Río, Ecuador |
|||||||||||||||||||||||||
|
Campos elevados |
f1 |
Pearsall 1987 |
Tipo de vestigio botánico: f=fitolito; g=grano de almidón; m=macro-vestigio; p=polen
Contexto: 1=sedimento arqueológico/campo agrícola; 2=artefacto
Tabla 1
Continuación
|
Sitio/Contexto |
Gossypium sp. (algodón) |
Malvaceae |
Solanaceae, cf. Capsicum spp. (ají) |
Solanaceae, cf. Capsicum frutenses (aji) |
Solanaceae, cf. Lycopersicum esculentum (tomate) |
Erythroxylaceae, cf. Erythroxylum coca (coca) |
Passifloraceae, cf. Passiflora nitida |
Asteraceae, cf. Ambrosia arborenscens (altamisa) |
Acanthaceae-Bignoniaceae, cf. Ruellia-Distictella |
Amarantaceae, cf. Gomphrena |
Caryophyllaceae, cf. Polycarpaea |
Chenopodiaceae, cf. Chenopodium quinoa (quinoa) |
Juglandaceae, pos Juglans |
Lythraceae, cf. Cuphea |
Moraceae, cf. Cecropia |
Myricaceae, cf. Myrica pubensces (palomero) |
Amaranthaceae, cf. Alternanthera |
Asteraceae |
Bromeliaceae, cf. Dyckia |
Anacardiaceae, cf. Pistacia |
Aristolochiaceae, cf. Aristolochia ringens (astroloja) |
Chrysobalanaceae, cf. Chrysobalanus icaco (icaco) |
Cyatheaceae, cf. Alsophyla |
Malvaceae, cf. Sida |
Myrtaceae, cf. Psidium guajava (guayaba) |
Onagraceae |
Plantaginaceae, cf. Plantago |
Polypodiaceae, cf. Polypodium |
Portulacaceae, cf. Talinum |
Symplocaceae, cf. Symplocos |
Trigonaceae, cf. Trigonia |
Fuente |
|
Llanos de Moxos, Bolivia Campos elevados El Villar, Santa Fe |
Erickson 1995 |
|||||||||||||||||||||||||||||||
|
Sitios habitación |
||||||||||||||||||||||||||||||||
|
Loma Mendoza |
Dickau et al. 2012 |
|||||||||||||||||||||||||||||||
|
Loma Salvatierra |
m1 |
m1 |
g2 |
Bruno 2010; Dickau et al. 2012 |
||||||||||||||||||||||||||||
|
Bella Vista |
Dickau et al. 2012 |
|||||||||||||||||||||||||||||||
|
Tumaco-Tolita, Colombia |
||||||||||||||||||||||||||||||||
|
Campos elevados |
||||||||||||||||||||||||||||||||
|
Las Tres Marias |
Patiño 2006 |
|||||||||||||||||||||||||||||||
|
Bajo Río San Jorge, Colombia |
||||||||||||||||||||||||||||||||
|
Sedimento lacunar |
p |
p |
p |
Herrera y Berrio 1998 |
||||||||||||||||||||||||||||
|
Peñon del Río, Ecuador |
||||||||||||||||||||||||||||||||
|
Campos elevados |
Pearsall 1987 |
Tipo de vestigio botánico: f=fitolito; g=grano de almidón; m=macro-vestigio; p=polen
Contexto: 1=sedimento arqueológico/campo agrícola; 2=artefacto
Tabla 1
Continuación
|
Sitio/Contexto |
Zea mays (maiz) |
Manihot esculenta (mandioca) |
Dioscoreaceae, Dioscorea spp. (ñame) |
Convolvulaceae, cf. Ipomoea (batata dulce) |
Araceae, cf. Xanthosoma sagittifolium (ocumo) |
Marantaceae |
Marantaceae, cf. Maranta arundinacea (arrurruz) |
Cannaceae, Canna edulis (achira) |
Heliconia sp. |
Cyperaceae |
Cucurbita spp. (zapallo) |
Cucurbitaceae, cf. Cucurbita maxima, C. mixta (zapallo) |
Arecaceae (palmeras) |
Arecaceae, cf. Acrocomia sclerocarpa (corozo) |
Astrocaryum standleyanum |
Phytelephas seemannii (tagua) |
Fabaceae |
Arachis hypogaeae (maní) |
Fabaceae, cf. Piptadenia peregrina (yopo) |
Fabaceae, cf. Herpalyce |
Fabaceae, cf. Indigofera tinctoria (añil) |
Aquifoliaceae, cf. Ilex paraguariensis (yerba mate) |
Bixaceae, cf. Bixa orellana (achiote) |
Amaranthaceae/Caryophyllaceae |
Fuente |
|
Río Daule, Ecuador Campos elevados |
f1 |
f1 |
f1 |
Stemper 1993 |
|||||||||||||||||||||
|
Costa de la Guyana Francesa |
|||||||||||||||||||||||||
|
Campos elevados |
|||||||||||||||||||||||||
|
Bajo Río Mana |
f1 |
f1 |
Iriarte et al. 2010 |
||||||||||||||||||||||
|
Grand Macuoa |
f1 |
Iriarte et al. 2010 |
|||||||||||||||||||||||
|
K-VIII |
f1 |
Iriarte et al. 2010 |
|||||||||||||||||||||||
|
Bois Diable |
f1 |
Iriarte et al. 2010 |
|||||||||||||||||||||||
|
Sitios habitación |
|||||||||||||||||||||||||
|
Sable Blanc |
g2 |
g2 |
Mckey et al. 2010 |
||||||||||||||||||||||
|
Barinas, Venezuela |
|||||||||||||||||||||||||
|
Campos elevados |
|||||||||||||||||||||||||
|
Caño Ventosidad |
p1 |
Salgado de Labouriau 1979 en Spencer 1994:131 |
|||||||||||||||||||||||
|
Campos drenados |
|||||||||||||||||||||||||
|
La Tigra, B27 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
Spencer et al. 1994 |
||||||||||||||||||
|
Sitio habitación |
|||||||||||||||||||||||||
|
La Tigra, B26 |
p1 |
p1 |
p1 |
p1 |
p1 |
Spencer et al. 1994 |
Contexto: 1=sedimento arqueológico/campo agrícola; 2=artefacto
Tabla 1
Continuación
|
Sitio/Contexto |
Gossypium sp. (algodón) |
Malvaceae |
Solanaceae, cf. Capsicum spp. (ají) |
Solanaceae, cf. Capsicum frutenses (aji) |
Solanaceae, cf. Lycopersicum esculentum (tomate) |
Erythroxylaceae, cf. Erythroxylum coca (coca) |
Passifloraceae, cf. Passiflora nitida |
Asteraceae, cf. Ambrosia arborenscens (altamisa) |
Acanthaceae-Bignoniaceae, cf. Ruellia-Distictella |
Amarantaceae, cf. Gomphrena |
Caryophyllaceae, cf. Polycarpaea |
Chenopodiaceae, cf. Chenopodium quinoa (quinoa) |
Juglandaceae, pos Juglans |
Lythraceae, cf. Cuphea |
Moraceae, cf. Cecropia |
Myricaceae, cf. Myrica pubensces (palomero) |
Amaranthaceae, cf. Alternanthera |
Asteraceae |
Bromeliaceae, cf. Dyckia |
Anacardiaceae, cf. Pistacia |
Aristolochiaceae, cf. Aristolochia ringens (astroloja) |
Chrysobalanaceae, cf. Chrysobalanus icaco (icaco) |
Cyatheaceae, cf. Alsophyla |
Malvaceae, cf. Sida |
Myrtaceae, cf. Psidium guajava (guayaba) |
Onagraceae |
Plantaginaceae, cf. Plantago |
Polypodiaceae, cf. Polypodium |
Portulacaceae, cf. Talinum |
Symplocaceae, cf. Symplocos |
Trigonaceae, cf. Trigonia |
Fuente |
|
Río Daule, Ecuador Campos elevados |
m1 |
Stemper 1993 |
||||||||||||||||||||||||||||||
|
Costa de la Guyana Francesa |
||||||||||||||||||||||||||||||||
|
Campos elevados |
||||||||||||||||||||||||||||||||
|
Bajo Río Mana |
Iriarte et al. 2010 |
|||||||||||||||||||||||||||||||
|
Grand Macuoa |
Iriarte et al. 2010 |
|||||||||||||||||||||||||||||||
|
K-VIII |
Iriarte et al. 2010 |
|||||||||||||||||||||||||||||||
|
Bois Diable |
Iriarte et al. 2010 |
|||||||||||||||||||||||||||||||
|
Sitios habitación |
||||||||||||||||||||||||||||||||
|
Sable Blanc |
g2 |
Mckey et al. 2010 |
||||||||||||||||||||||||||||||
|
Barinas, Venezuela |
||||||||||||||||||||||||||||||||
|
Campos elevados |
||||||||||||||||||||||||||||||||
|
Caño Ventosidad |
Salgado de Labouriau 1979 en Spencer 1994:131 |
|||||||||||||||||||||||||||||||
|
Campos drenados |
||||||||||||||||||||||||||||||||
|
La Tigra, B27 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
Spencer et al. 1994 |
||||||||||||||||||
|
Sitio habitación |
||||||||||||||||||||||||||||||||
|
La Tigra, B26 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
p1 |
Spencer et al. 1994 |
Tipo de vestigio botánico: f=fitolito; g=grano de almidón; m=macro-vestigio; p=polen
Contexto: 1=sedimento arqueológico/campo agrícola; 2=artefacto
La tabla 1 presenta diferentes tipos de vestigios botánicos de utilidad económica, los cuales han sido identificados a diferentes niveles taxonómicos. Ella comprende restos botánicos que han podido ser identificados a nivel de especie, como por ejemplo el maíz (Zea mays), y otras que han sido identificados a niveles taxonómicos más generales a nivel de género o familia, como por ejemplo las Marantáceas. Se ha marcado como “cf.” a aquellas plantas que solo han podido ser identificadas con seguridad a nivel de familia o genero y se ha sugerido de manera tentativa la posible especie de importancia económica que la misma puede estar representando. El lector deberá tener cuidado de distinguir claramente cuál es el nivel taxonómico especifico al cual un determinado vestigio botánico se ha identificado, ya que se corre el peligro de que esta interpretación de lo posible pasa a ser aceptado como un hecho en la literatura que surge posteriormente. En paréntesis se han colocado los nombres comunes de las plantas identificadas. Se debe también notar que la Tabla 1 solo refleja presencia/ausencia de los diferentes taxones de plantas en cada uno de los contextos. También se debe tener presente que los vestigios botánicos recuperados de los sitios de habitación y de los sedimentos lacustres, no necesariamente implica que los mismos hayan sido plantados en los campos elevados o drenados. Asimismo, por el momento, resulta arduo examinar con detalle la evolución histórica de estos sistemas agrícolas y delinear patrones temporales, ya que sin haber algunas excepciones, es difícil datar de manera precisa los campos elevados y drenados. La Figura 3 ilustra una selección de diferentes vestigios vegetales recuperados en sitios de agricultura de humedales y sitios habitacionales asociados.
La Tabla 1 muestra que una gran diversidad de plantas eran cultivadas en los campos agrícolas elevados y drenados, y en los sitios habitacionales asociados. Estos incluyen el maíz, tubérculos y raíces, vegetales como el zapallo y el maní, cultivos industriales como el algodón, tinturas como el añil y el achiote y hasta yerbas estimulantes y alucinógenas como la coca, la yerba mate, y el yopo.
El maíz es la planta domestica más ubicua en de los sistemas agrícolas de humedal pre-Colombinos. El mismo ha sido recuperado en todas las regiones de las tierras bajas de Sudamérica en donde se han realizado estudios arqueobotánicos. El maíz está representado en todas las clases de vestigios botánicos incluyendo los macro-vestigios botánicos, los fitolitos, los granos de almidón y el polen recuperados tanto de los rasgos agrícolas, los sedimentos lagunares, los sedimentos arqueológicos y en residuos de artefactos para procesar vegetales. Más aun, en algunos casos, como en los campos elevados de la Guyana Francesa, se han recuperado fitolitos diagnósticos de las hojas y de la mazorca del maíz, lo cual nos hace pensar que los residuos de la cosecha del maíz eran incorporados en los campos elevados y probablemente utilizados como abono verde (Iriarte et al. 2010).
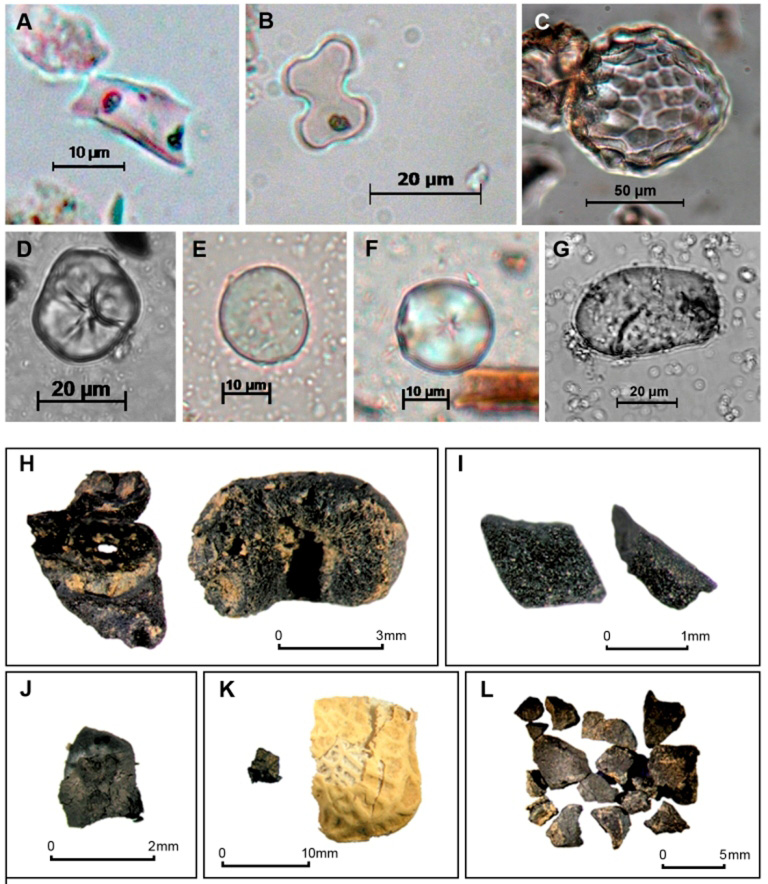
Figura 3 – Selección de vestigios botánicos recuperados en campos agrícolas elevados y sitios de habitación asociados. A. Fitolito de tipo “wavy-top rondel” de la mazorca de maíz (Zea mays) del sitio K-VIII, Guyana Francesa (tomado de Iriarte et al. 2010). B. Fitolito en forma de cruz Variedad 1 de la hoja de maíz (Zea mays) del sitio Bois Diable, Guyana Francesa (tomado de Iriarte et al. 2010). C. Fitolito de tipo “scalloped sphere” de la cascara de zapallo (Cucurbita spp.), Loma Salvatierra, Bolivia. D. Grano de almidón de maíz (Zea mays), Loma Salvatierra (tomado de Dickau et al. 2012). E. Grano de almidón de ají (cf. Capsicum sp.), Loma Salvatierra. F. Grano de almidón de mandioca (Manihot esculenta), Loma Salvatierra (tomado de Dickau et al. 2012). G. Grano de almidón de ñame (Dioscorea sp.), Loma Salvatierra (tomado de Dickau et al. 2012). H. Fragmento de mazorca y grano de maíz (Zea mays), Loma Salvatierra (tomado de Bruno 2010). I. Fragmento de cascara de zapallo (Cucurbita spp.), Loma Salvatierra (tomado de Bruno 2010). J. Base de semilla de algodón (Gossypium sp.), Loma Salvatierra (tomado de Bruno 2010). K. Fragmento de cascara arqueológica de maní, Loma Salvatierra (Arachis sp.) y moderna para comparación (tomado de Bruno 2010). L. Fragmentos de endocarpio de coquitos de palmera (Arecaceae) Loma Salvatierra (tomado de Bruno 2010).
Los tubérculos y raíces están menos presentes que el maíz. El que aparece en mayor cantidad de contextos es la mandioca, la cual aparece en los sitios habitacionales de los Llanos de Mojos, Beni (Dickau et al. 2012), de la Guyana Francesa (McKey et al. 2010) y en los sedimentos lacustres del Bajo Río San Jorge (Herrera y Berrio 1998). También se ha identificado el ñame (Dioscorea spp.) en los sitios habitacionales de Baures, Bolivia (Dickau et al. 2012), la batata (Ipomea batatas) en muestras de polen de un sitio habitacional en Barinas, Venezuela (Spencer et al. 1994) y se ha idenficado de manera tentativa el ocumo o la yalanga (Xanthosoma sagittifolium) en polen de campos elevados en el Beni, Bolivia (Erickson 1995) y en un sitio habitacional en Barinas, Venezuela (Spencer et al. 1994). La presencia de tubérculos pertenecientes a la familia de las Cannáceas como la achira (Canna edulis) y las Marantáceas como el arrurruz (Maranta arundinaceae) se han sugerido en los campos elevados del Río Daule, Ecuador (Stemper 1993). El polen y los fitolitos de las familias Cyperaceae y Heliconiaceae puede estar representando el cultivo y/o consumo de las raíces comestibles de algunas especies incluidas en estas familias (Dickau et al. 2012).
Entre otras plantas cultivadas se ha identificado el zapallo (Cucurbita spp.), el maní (Arachis hypogaeae) en los sitios habitacionales del Beni (Bruno 2010) y se ha sugerido el tomate (Solanaceae, cf. Lycopersicum esculentum) en los campos drenados de Barinas, Venezuela (Spencer et al. 1994). También se ha recuperado evidencia de frutas, como el polen de la familia Myrtaceae, el cual puede indicar el consumo de los frutos de la guayaba (cf. Psidium guajava), el icaco (Chrysobalanaceae, cf. Chrysobalanus icaco) en los sitios de habitación de Barinas, Venezuela (Spencer et al. 1994) y de la familia Passifloraceae que puede estar representando el maracuyá (cf. Passiflora nítida) en el bajo Río San Jorge (Herrera y Berrio 1998). Las palmeras están relativamente bien representadas en varios contextos habitacionales en el Beni, Bolivia (Bruno 2010; Dickau et al. 2012), en los campos de cultivo en Tumaco-Tolita en Colombia (Patiño 2006) y también en Barinas, Venezuela (Spencer et al. 1994).
Las condimentos como los ajíes (Capsicum spp.) están presentes en varios contextos asociados a la agricultura de humedales incluyendo sitios habitacionales en la costa de la Guyana Francesa (McKey et al. 2010) y el Beni (Dickau et al. 2012), los campos drenados de Barinas (Spencer et al. 1994) y sedimentos lacustres en el Bajo Río San Jorge (Herrera y Berrio 1998). Los cultivos industriales como el algodón (Gossypium sp.) se han recuperado de sitios de habitación del Beni, Bolivia (Bruno 2010), lo que concuerda con la presencia de gran cantidad de torteros en estos sitios de habitación (Jaimes Betancourt 2010). Se ha sugerido la presencia de tinturas como el añil (Fabaceae, cf. Indigofera tinctoria) en los sitios de habitación de Barinas (Spencer et al. 1994) y el achiote (Bixa sp.) en Bolivia (Erickson 1995). También se ha sugerido la presencia de yerbas medicinales, alucinógenas, estimulantes como el yopo (Fabaceae, cf. Piptadenia peregrina) tanto en los campos agrícolas drenados como sitios habitacionales de Barinas, Venezuela (Redmond y Spencer 2007), la coca (Erythroxylaceae, cf. Erythroxylum coca) en sedimentos lagunares del Bajo Río San Jorge (Herrera y Berrio 1998) y la yerba mate (Aquifoliaceae, cf. Ilex paraguariensis) en los campos elevados del Beni, Bolivia (Erickson 1995).
CONCLUSIONES
Si bien la arqueobotánica se encuentra aun en su infancia en las tierras bajas de Sudamérica, los datos resumidos brevemente en este artículo revelan patrones interesantes. Los datos indican que, sin lugar a dudas, el maíz fue el cultivo más ubicuo y probablemente uno de los cultivos más importantes que se plantaron en los campos elevados y drenados. Estos resultados son similares a los hallados en los estudios arqueobotánicos en la agricultura de humedales de Centroamérica que también indican que el maíz parece haber sido el cultivo predominante (p. e., Siemens et al. 1988; Turner y Miksicek 1984; Whitmore y Turner 2001). Nuevamente, debemos tener cuidado de no sobrevalorar los vestigios botánicos que tienden a preservar mejor como el maíz y las palmeras. Futuros estudios de isotopos estables de carbono en restos óseos humanos podrán brindar nuevos datos para esclarecer hasta qué punto el maíz fue el cultivo principal de la dieta de los diversos grupos de agricultores de humedales del Holoceno Tardío.
La presencia de maíz en todos los campos elevados de Sudamérica y Centroamérica no es algo novedoso. Sabemos que construir los campos elevados puede ser una tarea que precisa de gran cantidad de trabajo (p. e., Arco y Abrams 2006; Sanders et al. 1979). Asimismo, los estudios experimentales plantando maíz en campos elevados muestran que los mismos tienen la capacidad de alcanzar un gran rendimiento con una producción de hasta 5.8 t ha-1 (Arce 1993; Barba et al. 2003; Muse y Quintero 1987; Saavedra 2009; Sanders et al. 1979), lo cual es comparable a la alcanzada en los experimentos en Terra Preta Nova (Woods et al. en prensa).
También sabemos que la agricultura de humedales intensiva y de alta productividad del maíz sería capaz de proveer sustento para poblaciones numerosas. Como ilustramos en Iriarte et al. (2010: 2992) para una parcela de sabana inundable como Grand Macoua en la Guyana Francesa, que presenta 75 ha de superficie cultivable, tomando en cuenta a) la una estimación más baja obtenida de los experimentos de plantación de maíz en camellones modernos de 2 t ha-1 de maíz (Arce 1993), b) una sola cosecha anual, c) con solo un 25% de los campos elevados serian cultivados (dejando así tiempo para barbecho) y d) asumiendo un equivalente de 160kg de maíz de consumo anual por persona (el resto suplementado con otras actividades alimenticias) (Sanders et al. 1979), nos muestran que la productividad agrícola sería capaz de mantener a 234 personas. Una estimación moderada, asumiendo 75% de los campos elevados en uso y 3.725 t ha-1 (Sanders et al. 1979) de producción indica que el sitio podría haber sostenido 1310 personas. El máximo de personas que podrían haber sido mantenidas con una productividad de maíz de 5.78 t ha-1 (Muse y Quinteros 1987), 100% de los campos en uso y dos cosechas al año habría sido de 5400 personas. En base a estas estimaciones, utilizando un nivel de productividad y uso de la tierra moderado, podemos ver que la agricultura de maíz seria ciertamente capaz de mantener poblaciones numerosas.
Es así que resulta razonable pensar que estas poblaciones pre-Colombinas del Holoceno Tardío, no iban a invertir gran cantidad de trabajo en la construcción de campos agrícolas en humedales si no obtenían un cultivo de alto rendimiento como el del maíz. Aunque este es el tema de otro artículo en preparación, se debe notar que la agricultura de humedales intensiva de maíz parece estar conectada con las poblaciones numerosas y posiblemente jerárquicas que transformaron el paisaje a gran escala de las sabanas inundadas de las tierras bajas de Sudamérica durante el Holoceno Tardío (Erickson 2008; Gassón y Rey 2006; Lombardo y Prümers 2010; Plazas 1993; Prümers 2009; Rostain 2010b; Spencer y Redmond 1998; Walker 2004).
La mandioca y los demás tubérculos están menos representados que el maíz, pero se debe tener en cuenta que los tubérculos tienen problemas de preservación (Pearsall 1995) y en los contextos tropicales solo pueden ser recuperados a través de la extracción de residuos de los instrumentos que han sido utilizados para procesar vegetales en los sitios habitacionales y del polen de los sedimentos lagunares. Es así que se debe tener cuidado de no subestimar el rol de los tubérculos tropicales como la mandioca, la batata dulce o el ñame, entre otros, en relación a la preponderancia del maíz. Si bien el maíz aparece en todas las regiones, en todos los contextos y está representado en todas las clases de vestigios botánicos, el mismo no solo tiene mejores posibilidades de preservación debido a que es más robusto una vez carbonizado, sino que también el mismo posee la ventaja de poder ser identificado a nivel de especie por todos los tipos de vestigios botánicos como los macro-vestigios botánicos, los fitolitos, los granos de almidón y el polen. No se debe perder de vista que la mandioca también ha alcanzado altos rendimientos en los campos elevados alcanzando una producción de hasta 21 t ha-1 de mandioca (Arce 1993).
De la misma manera, algunos cultivos industriales como el algodón pueden haber sido importantes en la agricultura de humedales. Cabe señalar que los análisis de polen recuperados de los canales y los campos elevados de Albion Island también identificaron de manera tentativa la presencia de polen de posible algodón (Pulleston 1977). La presencia de macro-vestigios de algodón y numerosos fragmentos de torteros recuperados en las lomas de habitación de Bolivia apuntan a lo mismo (Bruno 2010; Dickau et al. 2012; Jaimes Bentancourt 2010).
Otro aspecto a destacar es que el análisis de los micro-vestigios botánicos recuperados en los instrumentos utilizados para procesar vegetales nos está aportando nueva información que nos está forzando a reevaluar algunas de las hipótesis sobre la utilización de los artefactos basados en su morfología y la analogía etnográfica. Este es el caso de la variedad de cultivos que han sido recuperados de los ralladores y coladores en las lomas de habitación de Salvatierra y Mendoza (Dickau et al. 2012), lo cual nos indica que estos instrumentos era utilizados para procesar varias plantas domesticas como el maíz y el ñame y no solamente la mandioca como ha sido sugerido tradicionalmente (p. e., ver Gongard 2006: 35).
En conclusión, como lo evidencia este breve resumen de los datos arqueobotánicos de los campos elevados y drenados de las tierras bajas de Sudamérica, los estudios arqueobotánicos nos están facilitando obtener evidencia directa de las plantas que fueron efectivamente cultivadas en estos rasgos agrícolas y consumidas en los sitios arqueológicos asociados, permitiendo superar años de especulaciones. Este resumen muestra una vez más la utilidad de combinar el estudio de macro- y micro-vestigios botánicos para documentar las plantas cultivadas en los ambientes húmedos de las tierras bajas sudamericanas revelando una gran diversidad de plantas, dentro de las cuales el maíz fue la planta domestica más ubicua y probablemente el cultivo predominante de la agricultura de humedales. En los próximos años, con la aplicación integrada y el continuo refinamiento de técnicas múltiples para la recuperación de vestigios vegetales, sin lugar a duda se expandirá la información relativa a la especies de importancia economía que fueron plantadas y consumidas por los agricultores de humedales de las tierras bajas de Sudamérica durante el Holoceno Tardío.
AGRADECIMIENTOS
Este trabajo fue producido bajo los auspicios Leverhulme Trust en el marco del proyecto “Impacto Humano y Uso de la Tierra en la Amazonia Boliviana” (F/00 158/CH). Mike Rouillard preparó las figuras del artículo.
REFERENCIAS
Alexandre, A., J.D. Meunier, A. Mariotti, y F. Soubies. 1999. Late Holocene phytolith and carbon-isotope record from a latosol at Salitre, South-Central Brazil. Quaternary Research 51: 187-194.
Arce, J. 1993. Evaluación y comparación de rendimientos de cuatro cultivos en tres anchuras de camellones (campos elevados) en la Estación Biológica del Beni (Prov. Ballivián, Dpto. Beni). Tesis de doctorado, Universidad Técnica del Beni, Trinidad, Bolivia.
Arco, L.J., y E.M. Abrams. 2006. An essay on energetics: the construction of the Aztec chinampa system. Antiquity 80: 906-918.
Barba, J., E. Canal, E. Garcia, E. Jorda, M. Miro, E. Pasto, R. Playa, I. Romero, M. Via, y E. Woynarovich. 2003. Moxos: Una Limnocultura. Cultura y medio ambiente en la Amazonia Boliviana. Barcelona: CEAM.
Bruno, M. C. 2010. Carbonized plant remains from Loma Salvatierra, Department of Beni, Bolivia. Zeitschrift für Archäologie Außereuropäischer Kulturen 3: 151-206.
Burn, M.J., y F.E. Mayle. 2008. Palynological differentiation between genera of the Moraceae family and implications for Amazonian palaeoecology. Review of Palaeobotany and Palynology 149: 187-201.
Bush, M. B., D. R. Piperno, P. A. Colinvaux, P. E. de Oliveira, L. A. Krissek, M. C. Miller, y W. E. Rowe. 1992. A 14,300-yr paleoecological profile of a lowland tropical lake in Panama. Ecological Monographs 62: 251-275.
Chandler-Ezell, K., D.M. Pearsall, y J.A. Zeidler. 2006. Root and tuber phytoliths and starch grains document manioc (Manihot esculenta) arrowroot (Maranta arundinacea) and llerén (Calathea sp.) at the real alto site Ecuador. Economic Botany 60: 103-120.
De las Casas, B. 1986 [1560]. Historia de las Indias (traducido del español). Caracas: Biblioteca Ayacucho.
Denevan, W.M. 1966. The Aboriginal Cultural Geography of the Llanos de Mojos of Bolivia. Berkeley: University of California Press.
____. 2001. Cultivated Landscapes of Native Amazonia and the Andes. New York: Oxford University Press.
____. 2007. Pre-European human impacts on tropical lowland environments, en The Physical Geography of South America. Editado por T.T. Veblen, K. R. Young y A.R. Orme, pp. 265-278. New York: Oxford University Press.
Denevan, W.M. Y B.L. Turner. 1974. Forms, functions and associations of raised fields in the old world tropics. Journal of Tropical Geography 39:24-33.
Denevan, W.M., y A. Zucchi. 1978. Ridged-field excavations in the central Orinoco Llanos, Venezuela, en In Advances in Andean Archaeology. Editado por D. L. Browman, pp. 235-245.The Hague: Mouton.
Denevan, W. M., D. Mathewson, y G. Knapp. 1987. Pre-Hispanic agricultural fields in the Andean region. Oxford: BAR International Series 359.
Denham, T.P., S.G. Haberle, C. Lentfer, R. Fullagar, J. Field, M. Therin, N. Porch, y B. Winsborough. 2003. Origins of agriculture at Kuk Swamp in the Highlands of New Guinea. Science 301: 189.
Dickau, R., A.J. Ranere, y R.G. Cooke. 2007. Starch grain evidence for the preceramic dispersals of maize and root crops into tropical dry and humid forests of Panama. Proceedings of the National Academy of Sciences EUA 104: 3651.
Dickau, R, M. C. Bruno, J. Iriarte, H. Prümers, C. Jaimes Betancourt, I. Holst, y F. E. Mayle. 2012. Diversity of cultivars and other plant resources used at habitation sites in the Llanos de Mojos, Beni, Bolivia: evidence from macrobotanical remains, starch grains, and phytoliths. Journal of Archaeological Science 39: 357-370.
Dillehay, T.D. 1997. Monte Verde: A Late Pleistocene Settlement in Chile, Volume II: The Archaeological Context. Washington DC: Smithsonian Institution Press.
Duncan, N.A., D.M. Pearsall, y R.A. Benfer. 2009. Gourd and squash artifacts yield starch grains of feasting foods from preceramic Peru. Proceedings of the National Academy of Sciences EUA 106: 13202.
Erickson, C.L. 1995. Archaeological methods for the study of ancient landscapes of the Llanos de Mojos in the Bolivian Amazon, en Archaeology in the Lowland American Tropics. Editado por P. Stahl, pp. 66-95. Cambridge: Cambridge University Press.
____. 2008. Amazonia: the historical ecology of a domesticated landscape, en The Handbook of South American Archaeology. Editado por H. Silverman and W. Isbell, pp. 157-183. New York: Springer.
Gassón, R., y J.C. Rey. 2006. Cacicazgos ciclicos e intensificacion agricola en los Llanos Occidentales de Venezuela, en Agricultura Ancestral. Camellones y Albarradas. Contexto Social, Usos y Retos del Pasado y del Presente. Editado por F. Valdéz, pp. 141-158. Ecuador: Abya Yala.
Gongard, P. 2006. Campos elevados en llanuras humedas del modelado al paisaje camellones, waru warus o pijales, en Agricultura ancestral. Camellones y albarradas. Contexto social, usos y retos del pasado y del presente. Editado por F. Valdéz, pp. 25-53. Ecuador: Abya Yala.
Gumilla, J. 1963 [1791]. The Orinoco illustrated and defended (Traducido del español). Caracas: Biblioteca de la Academia Nacional de la Historia.
Heckenberger, M., y E.G. Neves. 2009. Amazonian archaeology. Annual Review of Anthropology 38: 251-266.
Herrera, L. F., y L.E Urrego. 1996. Atlas de polen de plantas utiles y cultivadas de la Amazonia Colombiana. Bogotá: Fundación Erigaie.
Herrera, L.E., y J.C. Berrio. 1998. Vegetación natural y acción antrópica de los últimos 1000 años en el sistema prehispánico de canales artificiales del Caño Carate en San Marcos (Sucre, Colombia). Corpoica. Revista de la Corporación Colombiana de Investigación Agropecuaria 2: 2-35.
Hurd, P.D., E.G. Linsley, y T.W. Whitaker. 1971. Squash and gourd bees (Peponapis, Xenoglossa) and the origin of the cultivated Cucurbita. Evolution 25 (1): 218-234.
Iriarte, J. 2003. Mid-Holocene emergent complexity and landscape transformation: the social construction of early formative communities in Uruguay, La Plata Basin. Tesis de Doctorado, Departamento de Antropología, University of Kentucky, EUA.
____. 2006. Vegetation and climate change since 14,810 C-14 yr BP in southeastern Uruguay and implications for the rise of early Formative societies. Quaternary Research 65: 20-32.
Iriarte, J., I. Holst, O. Marozzi, C. Listopad, E. Alonso, A. Rinderknecht, y J. Montaña. 2004. Evidence for cultivar adoption and emerging complexity during the mid-Holocene in the La Plata basin. Nature 432: 614-617.
Iriarte, J., B. Glaser, J. Watling, A. Wainwright, J. J. Birk, D. Renard, S.Rostain, y D. McKey. 2010. Late Holocene Neotropical agricultural landscapes: phytolith and stable carbon isotope analysis of raised fields from French Guianan coastal savannahs. Journal of Archaeological Science 37: 2984-2994.
Iriarte, J., Power, M., Mayle, F., Rostain, S., Jones, H., Watling, J., Brownen, W., McKey, D. 2012. Fire-free land use in pre-1492 Amazonian savannas. Proceedings of the National Academy of Sciences 19:6473-6479.
Jaimes Betancourt, C. 2010. La cerámica de la Loma Salvatierra. Tesis de Doctorado, Universidad de Bonn, Alemania.
Jarosz, N., B. Loubet, B. Durand, A. Mccartney, X. Foueillassar, y L. Huber. 2003. Field measurements of airborne concentration and deposition rate of maize pollen. Agricultural and Forest Meteorology 119: 37-51.
Kealhofer, L., y D. Penny. 1998. A combined pollen and phytolith record for fourteen thousand years of vegetation change in northeastern Thailand. Review of Palaeobotany and Palynology 103:83-93.
Lombardo, U. 2010. Raised fields of northwestern Bolivia: a GIS based analysis. Zeitschrift für Archäologie Außereuropäischer Kulturen 3: 127-149.
Lombardo, U., E. Canal-Beeby, S. Fehr, y H. Veit. 2011. Raised fields in the Bolivian Amazonia: a prehistoric green revolution or a flood risk mitigation strategy? Journal of Archaeological Science 38: 502-512.
Lombardo, U., y H. Prümers. 2010. Pre-Columbian human occupation patterns in the eastern plains of the Llanos de Moxos, Bolivian Amazonia. Journal of Archaeological Science 37: 1875-1885.
McKey, D., S. Rostain, J. Iriarte, B. Glaser, J. J. Birk, I. Holst, y D. Renard. 2010. Pre-Columbian agricultural landscapes, ecosystem engineers, and self-organized patchiness in Amazonia. Proceedings of the National Academy of Sciences 107: 7823-7828.
Mulholland, S.C. 1993. A test of phytolith analysis at Big Hidatsa, North Dakota, en Current Research in Phytolith Analysis: Applications in Archaeology and Paleoecology. Editado por D.M. Pearsall y D.R. Piperno, pp. 131-145. Philadelphia: University of Pennsylvania, MASCA Research Papers in Science and Archaeology.
Muse, M., y F. Quintero. 1987. Experimentos de reactivación de campos elevados, Peñón del Río, Guayas, Ecuador, en Pre-Hispanic Agricultural Fields in the Andean Region. Editado por W.M. Denevan, D. Mathewson y G. Knapp, pp. 249-266. Oxford: BAR International Series 359.
Parsons, J.J., y R. Shlemon. 1987. Mapping and dating the prehistoric raised fields of the Guayas basin, Ecuador, en Pre-Hispanic Agricultural Fields in the Andean Region. Editado por W.M. Denevan, D. Mathewson y G. Knapp, pp. 207-216. Oxford: BAR International Series 359.
Patiño, C. 2006. Campos prehispánicos elevados en la economía Tumaco-Tolita, en Agricultura ancestral. Camellones y albarradas. Contexto social, usos y retos del pasado y del presente. Editado por F. Valdéz, pp. 169-188. Ecuador: Abya Yala.
Pearsall, D.M. 1987. Evidence for prehispanic maize cultivation on raised fields at the Peñon del Río, Guayas, Ecuador, en Pre-Hispanic Agricultural Fields in the Andean Region. Editado por W.M. Denevan, D. Mathewson y G. Knapp, pp. 279-295. Oxford: BAR International Series 359.
_____. 1995. “Doing” paleoethnobotany in the tropical lowlands: adaptation and innovation in methodology, en Archaeology in the Lowland American Tropics: Current Analytical Methods and Recent Applications. Editado por P. Stahl, pp. 113-129. Cambridge: Cambridge University Press.
Pearsall, D.M., K. Chandler-Ezell, y A. Chandler-Ezell. 2003. Identifying maize in neotropical sediments and soils using cob phytoliths. Journal of Archaeological Science 30: 611-627.
Perry, L. 2004. Starch analyses reveal the relationship between tool type and function: an example from the Orinoco valley of Venezuela. Journal of Archaeological Science 31: 1069-1081.
_____. 2005. Reassessing the Traditional Interpretation of “Manioc” Artifacts in the Orinoco Valley of Venezuela. Latin American Antiquity: 409-426.
Piperno, D.R. 2006. Phytoliths: A Comprehensive guide for archaeologists and paleoecologists. San Diego: Altamira Press.
Piperno, D.R., y J.G. Jones. 2003. Paleoecological and archaeological implications of a Late Pleistocene/Early Holocene record of vegetation and climate from the Pacific coastal plain of Panama. Quaternary Research 59: 79-87.
Piperno, D.R., y K.E. Stothert. 2003. Phytolith evidence for early Holocene Cucurbita domestication in southwest Ecuador. Science 299: 1054.
Piperno, D.R., A.J. Ranere, I. Holst, and P. Hansell. 2000. Starch grains reveal early root crop horticulture in the Panamanian tropical forest. Nature 407: 894-897.
Plazas, C. 1993. La sociedad hidráulica Zenú: Estudio arqueológico de 2.000 años de historia en las llanuras del Caribe Colombiano. Bogotá: Banco de la República, Museo del Oro.
Plazas, C., y A. M. Falchetti. 1987. Poblamiento y adecuación hidráulica en el bajo río San Jorge, costa Atlántica, Colombia, en Pre-Hispanic agricultural fields in the Andean Region. Editado por W.M. Denevan, D. Mathewson y G. Knapp, pp. 483-503. Oxford: BAR International Series 359.
Prümers, H. 2009. “Charlatanocracia” en Mojos? Investigaciones arqueológicas en la Loma Salvatierra. Beni, Bolivia. Boletín de Arqueología PUCP 11: 103-116.
____. 2010. Die Untersuchungen der Jahre 2007-2008 zur Vorspanischen Siedlungsgeschichte in den Llanos de Mojos (Bolivien). Zeitschrift für Archäologie Außereuropäischer Kulturen 3: 233-244.
Pulleston, D.E. 1977. The art and archaeology of hydraulic agriculture in the Maya Lowlands, en Social Process in Maya Prehistory. Editado por N. Hammond, pp. 449-467. New York: Academic Press.
Real, L.A. 1981. Nectar availability and bee-foraging on Ipomoea (Convolvulaceae). Biotropica: 64-69.
Redmond, E. M., y C. Spencer. 2007. Archaeological Survey in the High Llanos and Andean Piedmont of Barinas, Venezuela. Vol. 86. New York: Anthropological Papers of the American Museum of Natural History.
Renard, D., J.J. Birk, B. Glaser, J. Iriarte, G. Grisard, J. Karl, y D. McKey. En prensa a. Origin of mound-field landscapes: a multi-proxy approach combining contemporary vegetation, carbon stable isotopes and phytoliths. Plant and Soil. doi: 10.1007/s11104-011-0967-8.
Renard, D., J. Iriarte, J.J. Birk, S. Rostain, B. Glaser, y D. McKey. 2011. En prensa b. Ecological engineers ahead of their time: The functioning of pre-Columbian raised-field agriculture and its potential contributions to sustainability today. Ecological Engineering. doi:10.1016/j.ecoleng.2011.03.007.
Rival, L., y D. McKey. 2008. Domestication and diversity in manioc (Manihot esculenta Crantz ssp. esculenta, Euphorbiaceae). Current Anthropology 49: 1119-1128.
Rogers, D.J. 1965. Some botanical and ethnological considerations of Manihot esculenta. Economic Botany 19: 369-377.
Rostain, S. 2008. Agricultural earthworks on the French Guiana coast, en The Handbook of South American Archaeology. Editado por H. Silverman and W. Isbell, pp. 217-233. New York: Springer.
____. 2010a. Pre-Columbian earthworks in coastal Amazonia. Diversity 2: 331-352.
____. 2010b. Cacicazgos guyanenses: mito o realidad?, en Arqueologia Amazônica. Vol. I. Editado por E. Pereira y V. Guapindaia, pp. 167-192. Belém: MPEG/IPHAN.
Saavedra, O. 2009. Culturas hidráulicas de la Amazonia Boliviana. La Paz: OXFAM.
Sanders, W.T., J.R. Parsons, y R.S. Santley. 1979. The basin of Mexico: Ecological processes in the evolution of a civilization. New York: Academic Press.
Siemens, A.H., R.J. Hebda, M.N. Hernández, D.R. Piperno, J.K. Stein, y M.G. Zolá Báez. 1988. Evidence for a cultivar and a chronology from patterned wetlands in central Veracruz, Mexico. Science 242: 105.
Sneider Rojas, M., and F. G. Montejo. 2006. Manejo deI espacio y aprovechamiento de recursos en la depresión Momposina, Bajo Río San Jorge, en Agricultura Ancestral. Camellones y albarradas. Contexto social, usos y retos del pasado y del presente. Editado por F. Valdéz, pp. 81-91. Ecuador: Abya Yala.
Spencer, C.S., y E.M. Redmond. 1998. Prehispanic causeways and regional politics in the llanos of Barinas, Venezuela. Latin American Antiquity 9(2):95-110.
Spencer, C. S., E. M. Redmond, y M. Rinaldi. 1994. Drained fields at la Tigra, Venezuelan Llanos: a regional perspective. Latin American Antiquity 5(2): 119-143.
Stemper, D.M. 1993. The persistence of prehispanic chiefdoms on the río Daule, Coastal Ecuador. University of Pittsburgh Memoirs in Latin American Archaeology. Pittsburgh: University of Pittsburgh.
Turner, B. L. II, y W. Denevan. 1985. Prehistoric manipulation of wetlands in the Americas: A raised field perspective, en Prehistoric Intensive Agriculture in the Tropics. Editado por I. Farrington, pp. 11-30. Oxford: BAR International Series 232.
Turner, B.L., y P.D. Harrison. 1983. Pulltrouser swamp: Ancient Maya habitat, agriculture, and settlement in northern Belize. Austin: University of Texas Press.
Turner, B.L., y C.H. Miksicek. 1984. Economic plant species associated with prehistoric agriculture in the Maya lowlands. Economic Botany 38(2): 179-193.
Ugent, D., S. Pozorski, y T. Pozorski. 1986. Archaeological manioc (Manihot) from coastal Peru. Economic Botany 40(1): 78-102.
Valdéz, F. 2006. Agricultura Ancestral Camellones y Albarradas: Contexto Social, Usos y Retos del Pasado y del Presente: Coloquio Agricultura Prehispánica Sistemas Basados en el Drenaje y en la Elevación de los Suelos Cultivados. Ecuador: Editorial Abya Yala.
Versteeg, A.H. 2003. Suriname before Columbus. Paranaibo: Stichting Surinaams Museum.
Walker, J. H. 2004. Agricultural Change in the Bolivian Amazon. Pittsburgh: University of Pittsburgh Memoirs in Latin American Archaelogy 13.
Whitmore, T.M., y B.L. II Turner. 2001. Cultivated Landscapes of Middle America on the Eve of Conquest. Oxford: Oxford University Press.
Whitney, B. S., A. C. Rushton, J.F. Carson, J. Iriarte, and F. Mayle. En prensa. An improved methodology for the recovery of Zea mays and other large crop pollen, with implications for environmental archaeology in the Neotropics. The Holocene.
Woods, W.I., W. M. Denevan, y L. Rebellato. En prensa. Population estimates for anthropogenically enriched soils (Amazonian Dark Earths), en Living on the Land: the Complex Relationship Between Population and Agriculture in the Americas. Editado por J. Wingard y S. Hayes. Boulder: University of Colorado Press.
Recebido em 15/02/2012.
Aprovado em 08/03/2012.
Amazônica 4 (1): 30-58, 2012